Клас Плазуни (Reptilia)
РЯД Лускаті (Squamata)
Родина Веретільницеві (Anguidae)
Веретільниця східна (колхидська) чи ламка? (Anguis fragilis? Anguis colchica?)
Охоронний статус
Відсутній
Опис зовнішнього вигляду
Порівняно велика безнога ящірка зі змієподібним тілом загальною довжиною до 50 см. Самці трохи менші за самиць. Хвіст відносно короткий (не довший за тулуб), дуже ламкий. Вушний отвір менший або рівний ніздрям. Луска тулуба гладенька, розташована у поздовжні рядки. Зуби маленькі і гострі. Візуально два близькі види, котрі до останнього часу були підвидами, дуже складно відрізняються. На відміну від отруйної гадюки у веретільниці нема звуження між головою і тілом. Шия у безногої ящірки товста і у неї не помітний перехід від голови до тіла. Наявність пістрявого забарвлення у самиць веретільниці, мідянки, гадюки створює для новачків проблеми у визначенні цих видів за кольором.
Рис. 1. Зовнішній вигляд самця веретільниці (Фото С. Жили).

Молоді веретільниці зверху сріблясто-білого або блідо-кремового кольору з двома тісно розташованими темними смугами, які тягнуться від розташованої на потилиці трикутної плямки. Боки й черево чорного або чорно-бурого забарвлення, причому межа між світлою спинною і темної бічною частиною тіла виражена дуже різко. У дорослих ящірок спинна сторона тулуба поступово темнішає і набуває коричнево-бурого або темно-сірого кольору з характерним бронзовим відливом. За цей коричнево-бронзовий колір веретільницю цілком закономірно звуть у народі «мідянка-мідяниця». Довжина тіла з хвостом 23-43 см, маса 15-35 г. Довжина тіла веретільниць з території Білорусі становить 11,5-21,2 см (♂ - 11,5-17,4, ♀ - 12,4-21,2 см), довжина хвоста -11,6-20,6 см (♂ - 11,6-17,0, ♀ - 13,2-20,6 см), довжина голови - 1,1-1,5 см. Довжина тіла поліських екземплярів виявляється дещо меншою, ніж максимальна для ареалу в цілому - 265 мм (Пікулік, 1996).
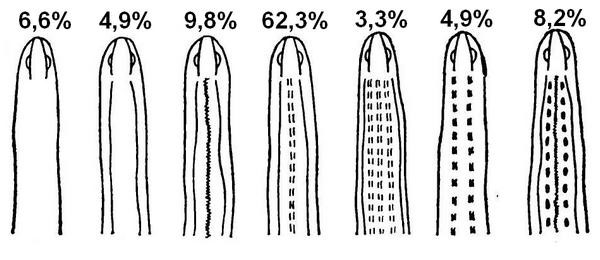
Рис. 2. Комбінація форм візерунку верхньої частини тіла веретільниць з білорусі (Пикулик, 1988).

Поширення - вся територія Полісся. Місцеві назви - мідяниця, слівень, мядзянка, мядзяніца. У Європі цей вид можуть називати «повільним хробаком». Єдиний представник безногих ящірок у Поліссі. Повзає повільно, роблячи характерні незграбні широкі і нерівні рухи. На відміну від інших ящірок Білорусі веретінниця мало помітна в природі. Веде досить прихований спосіб життя. Цю змію рідко зустрічають у природі і часто навіть не здогадуються про її існування. До того ж вона активна в густій рослинності, основному в сутінках і вночі при теплій погоді. Веретільницю місцеве населення помилково вважає «отруйною мідянкою» і безжально знищує. Найбільше виявляється і гине веретільниць під час збору чорниці. Вдень активна в похмуру погоду, хоча відзначалися окремі випадки активності веретільницю в спекотну погоду на піску при температурі понад 30 ° C. Цих безногих ящірок, які гріються на сонці, частіше можна бачити навесні, коли ще недостатньо тепла, а також після періоду холодної погоди влітку. Любить ця ящірка вийти на полювання після літнього дощу. Однак під час подолання піщаної дороги рухається вкрай повільно і тільки у заростях трави, чагарнику, на камінні її рухи стають більш вправними енергійними і швидкість руху зростає. Полює ця ящірка на дощових черв'яків, яких буває багато на поверхні ґрунту після дощу. Незважаючи на змієподібний вигляд веретільниця - це безнога ящірка. Всі ящірки відрізняються від змій за «видимими» повіками, а не «скляними» та здатністю відкидати хвіст. Самиці, як правило, мають темні боки і тонку, темну смужку. Вони також мають порівняно менші голови, ніж самці. Самці переважно мають однорідний сірий колір, позбавлений поздовжньої смуги та часто розсіяні по тілу сині плямки. Старі веретільниці можуть мати шрами від поєдинків. У тих місцях, де ви зустріли таку мідянку, можна у кінці травня спробувати поспостерігати за шлюбними турнірами цього виду. Самиці більші, але самці мають більший відносний розмір голови. При однаковій швидкості росту у самців відзначається непропорційне збільшення розміру голови, а у самиць відповідно розміру тіла. Ця особливість поряд з деякими іншими корисна при визначенні статі. У цих тварин різниця у відносно більшому розмірі голови у самців збільшується з віком. Ці статеві відмінності добре пояснюються репродуктивною біологією виду, а саме видовищними і енергійно затратними шлюбними турнірами, наявністю копулятивних укусів у великих самиць. Великі розміри порівняно довгих самиць можна розглядати, як доказ того, що їх плодючість залежить від величини. Відмінність у розмірах зумовлює і дивергенцію у харчуванні між статями. Однак відносно великий розмір голови самців веретільниць навряд чи може розглядатись як якась серйозна зброя у добуванні жертв (Tibor S., Gábor H., 2009). З позицією Tibor, Gábor (2009) не можна погодитись з тої причини, бо різнорозмірність тіла, форма голови веретільниць мають велике і різнофункціональне адаптивне пристосування, котре дає змогу цій ніби то примітивній і незграбній ящірці бути конкурентоздатною у дикій природі поряд з сильними вужами, гадюками та мідянками.
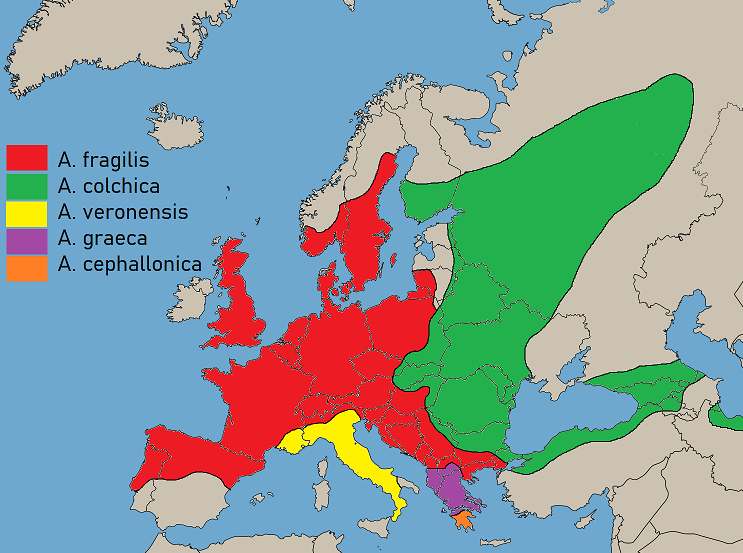
Європа нині переживає небувалий сплеск генетичних досліджень для більшості видів диких тварин. Такі роботи проводяться і для веретільниць. Так після проведення аналізу мітохондріальної ДНК роду Anguis для переважної більшості Європи, стало зрозумілим, що у нас в Україні поширена скоріше не веретільниця ламка, а зовсім інший вид Anguis colchica, котрий раніше був підвидом (Anguis fragilis colchica). Види-двійники – явище досить поширене у природі. Без проведення генетичних досліджень саме української популяції Anguis у повному обсязі не можна говорити про встановлену видову приналежність нашої веретільниці. На сьогодні для офіційного визнання українського виду як Anguis colchica відсутні необхідні наукові підстави. Філогенетичні взаємозв'язки роду безногих ящірок Західної Палеарктики були зроблені на основі аналізу фрагменту мітохондріальної ДНК та двох локусів, що кодують ядерний білок. Веретільниця «колхіка» має великий ареал від сходу Чехії та Балтійського регіону і на схід включно північний Іран. Цей вид ймовірно зустрічається у східній Скандинавії та на північно-східних Балканах (Gvozˇdíka, 2010). Філогенетична класифікація роду Anguis зазнала глибоких змін за останні роки і ці зміни ще не зовсім визнані в «українській систематиці» і відповідно у назві цього виду веретільниці. Хоча за зовнішніми ознаками (морфологічно) види роду Anguis майже однакові, але вони із західної Палеарктики були генетично ідентифіковані як чотири самостійні і широкопоширені види: A. fragilis Linnaeus, 1758, A. graeca Bedriaga, 1881, A. colchica (Nordmann, 1840) та A. cephallonica Werner, 1894 (Gvoždík et al., 2010). Існуюча закономірність генетичної диференціації, ймовірно, була наслідком географічних бар'єрів (головним чином гірських систем), що призвело до ізоляції алопатричних популяцій в районі, відповідно до моделі «рефугії» чи «локалітету» (Jablonski et al., 2016). Однак західна межа A. сolchica у Європі лише частково може бути пояснена наявністю географічного бар’єру, бо гори Карпати потрапили всередину ареалу східної веретільниці.
Рис. 3. Поширення східної веретільниці у Європі (зелений колір).

Опис екології виду
Сучасна структура поліської популяції веретільниці є результатом процесів глобального потепління клімату, історичних змін ландшафтів, осушувальних робіт, способів господарювання та наявності у минулому лісового випасу худоби, котрий був насправді сприятливим фактором для формування життєздатних локалітетів цього виду. Оселища цього виду потребують наявності достатнього шару лісової підстилки чи трав’яного очісу у поєднанні з місцями, де можна погрітись, з розрідженими та освітленими сонцем полянками лісів Оселища залежать і від технологій проведення рубок. Для безногої ящірки сприятливі зруби малої площі. Очевидно на генетичне різноманіття поліської популяції цього виду матиме вплив скорочення чи подекуди розширення існуючих мікролокалітетів ареалу, створення перешкод-бар’єрів, до яких цей порівняно повільний і незграбний вид дуже чутливий Тому в Україні потрібні генетичні дослідження рептилій. Полісся і у т. ч. ЧРЕБЗ є одним з найважливіших регіонів європейського біорізноманіття, де багато видів помірного клімату збереглося з часів закінчення плейстоценових заледенінь і де висока топографічна неоднорідність забезпечує належні умови для місцевої адаптації популяцій. Вивчення особливостей харчування веретільниці проводиться переважно за методикою аналізу вмісту шлунку та кишковика на особинах, знайдених мертвими в природі. Веретільниці полюють на безліч безхребетних, в основному на земляних червів (33,33% вмісту шлунку добутих особин) та равликів-слимаків (35,41%). Спектр живлення A. Fragilis чи A. сolchica у Поліссі, ймовірно, залежить від характеру активності цієї ящірки, яка активна переважно після дощу, у перші світлі години на світанку та в сутінках.
Для веретільниці характерні ізольовані популяції у Італії та західній Скандинавії. Цей вид більшу частину свого життя проводить під землею і його цілком закономірно відносять до напівфосоріальних. Польові дослідження у Поліссі вказують на існування локалітетів з більш високими чисельностями цього виду у напіввідкритих лісових біотопах (структурованих лісах з полянами, сіножатями, випасом худоби, існуючою системою весняних випалів невеликих площ). Саме ці біотопи у Поліссі зазнали інтенсивної трансформації і у наш час веретільниця часто зустрічається на околицях лісових сіл з невеликою чисельністю населення, на перелогах городів, котрі використовуються під випас малочисельної худоби. Як і у минулому ця безнога ящірка чисельна в екотонах різних типів лісу – сухого та вологого з наявністю освітлених сонцем піщаних оголень ґрунту, де рептилії можуть погрітись. В умовах ЧРЕБЗ біотопи або оселища цього виду знаходяться у деградованому стані.
Фенологія виду і особливості розмноження у Південній, Західній і Центральній Європі добре вивчені. Так був простежений репродуктивний цикл 68 пологів та 468 новонароджених. Встановлені основні репродуктивні характеристики досліджуваних популяцій Європи, такі як розмір виплоду, вага потомства, розмір потомства та мінімальний розмір зрілості для самок: довжина морди 135-150 мм. Взаємозв'язок між величиною приплоду та розмірами самиць веретільниць. Очевидно, що ця закономірність характерна і для інших видів ящірок та змій. Для півдня Європи характерна більш висока інтенсивність розмноження, чим для Полісся. Там 88,8% дорослих самиць розмножуються щорічно (Galán, 2004). У Поліссі цей вид приносять приплід, як правило, через рік і можливо переважно на південно-західній частині ареалу України щорічно. Веретільниця не є строго територіальним видом і агресивні стосунки характерні виключно для періоду розмноження. Вагітні самиці мають дещо іншу поведінку і їм очевидно потрібно більше сонячного світла на обігрів тіла, щоби прискорити розвиток яєць. Облаштування штучних надійних схованок у перспективних місцях для прийняття сонячних ванн та високими кормовими умовами може бути дієвим заходом охорони не тільки веретільниці, але і інших рептилій. Польові спостереження в умовах ЧРЕБЗ вказують на те, що добрим замінником оголеного піщаного ґрунту може бути шматок шиферу, котрий теж як і відкритий пісок дуже швидко нагрівається сонцем.
Останнім часом чисельність у Європі скорочується з причини руйнування і фрагментації її оселищ.
Веретільниця – це гарний приклад адаптації виду до життя у нірках, під купами рослинного вітошу. Цю тендітну ящірку не можна відловлювати, затискати, бо вона у цьому разі позбудеться хвоста. Полишення хвоста під час небезпеки - це захисна відповідь, яка відволікає хижака і допомагає врятуватися. Самокалічення шляхом відкидання якоїсь частини тіла називається автотомією. Серед хребетних тварин її застосовують тільки ящірки. У випадку небезпеки, кільцеві м’язи на хвості різко скорочуються, розриваються і хвіст відлітає в бік. Оскільки нервові закінчення у відірваному хвості продовжують ще деякий час функціонувати, вони змушують м’язи мимовільно скорочуватись. Частина відокремленого хвоста часто з більш яскравим забарвленням починає швидко згинатися, стрибати.
Для веретільниці характерний канібалізм, але очевидно його значення в екології і динаміці чисельності цього виду є перебільшеним. Можна припустити, що ці ящірки просто не роблять чи й не мають змоги визначати видову приналежність жертви, котру потрібно швидко схопити. Канібалізм - це механізм, який підвищує успішність виживання особин і спостерігається від найпростіших до ссавців (Polis 1981, Polis & Myer 1985). Він також відносно поширений у земноводних та плазунів (Polis 1981, Polis & Myers 1985, Mitchell 1986). Канібалізм характеризується чотирма аспектами: вбивство жертв, отримання енергії від жертв, взаємодія залежно від розміру та внутрішньовидова конкуренція (Claessen et al. 2004). Загалом, канібалізм виглядає генетично, але контролюється або викликається різними екологічними ознаками (Polis 1981, Polis & Myers 1985). Голодування, екологічний та харчовий стрес можуть стимулювати канібалізм (Polis 1981). Як правило, канібалістичні особини можуть бути не в змозі виявити особини візуально чи хімічно через фізіологічні чи неврологічні дефекти, і у таких осіб візуальні або хімічні подразники можуть спричинити хижацьку реакцію (Mitchell 1986). У більшості видів плазунів канібалізм виявляється опортуністично як побічний продукт нормальної хижацької поведінки (Polis & Myer 1985).
Вид населяє ліси, луки, сади та компостні відвали (Başoğlu & Baran 1977, HCT 2010). Харчування повільного хробака складається з різноманітних безхребетних, особливо земляних черв'яків та слимаків (Luiselli 1992, Luiselli et al. 1994, Pedersen et al. 2009, Mollov 2010). У 2010 року, дорослий самець (168 мм) з’їв неповнолітнього (75 мм), котрий вкусив за голову. Раніше про канібалізм у Anguis fragilis повідомляли Деліц (1921), Петцольд (1971), Девіс (1967) та Фрейті (1987). Пецольд (1971) зазначив, що неповнолітні, які перебувають у полоні, їдять один одного. Девіс (1967) повідомив про самицю, яка з’їла двох своїх новонароджених нащадків у неволі. Fretey (1987) згадував, що A. fragilis іноді може полювати на дрібних ящірок (L. vivipara, A. fragilis) та неповнолітніх змій (N. natrix).
Неповнолітніх особин, як правило, їдять дорослі особини багатьох видів ящірок (Polis & Myers 1985, Mitchell 1986). Більше того Мітчелл (1986) заявив, що наявність відмінностей у кольорі та малюнку між дорослими та неповнолітніми може бути результатом природного відбору для уникнення канібалізму. Він також повідомив, що частота неповнолітніх канібалізму може бути пов'язана з щільністю населення в природних популяціях. Однак причиною цієї справи може бути екологічний (наприклад, простір, температура, освітленість) та харчовий стрес (наприклад, голод, переваги їжі) у неволі. Умови неволі не могли повністю задовольнити фізіологічні потреби організму, збереження здоров’я, поведінки та психології виду. За Купером (1990), веретільниці можуть розрізняти хімічні подразники, що виникають від здобичі, та інші пахучі подразники, як інші активні у полюванні види ящірок, а їх язик - це орган, який збирає молекули для аналізу за допомогою вомероназального органу.
Поведінка канібалізму у активного фуражеру під умовною назвою «Повільний черв’як» (Ghira et al. 1999) пояснювала ефективну техніку добування їжі за принципом «сиди і чекай». Для цього не потрібно щось особливе робити, щоб отримати їжу і не робити повторні відбори проб жертв у місці засідки. Більше того, виявлення здобичі помахуванням якоїсь частини тіла, як наприклад гримучого хвоста, може бути замінене нерухомістю (Купер 1997). Адаптація добування їжі з засідки може спричинити втрату необхідності хімічного визначення здобичі (Cooper & van Wyk, 1994). Оскільки ящірки перебувають у неволі, нерухомість може призвести до того, що доросла особина неправомірно визначить неповнолітнього повільного хробака як здобич. Поліс (1981) повідомив, що у 86% із 50 випадків, коли були зафіксовані статеві відмінності у хижака, самиці були у більшій мірі канібалами. Подальші спостереження не підтвердило цього твердження. Більшість даних про канібалізм є безперечно неповними, не завжди обґрунтованими і не дають уявлення про причини та наслідки такої поведінки у веретільниць (Mitchell 1986). Для отримання детальних даних про канібалізм та розуміння його механізму слід здійснити майбутні комплексні роботи і не можна поведінку виду в умовах неволі прирівнювати до його дій у природі.
Веретільниці зрідка з'їдаються зміями (мідянкою і гадюкою). Вуж і безнога ящірка у більшій мірі розділені біотопічно. Наявність порівняно великої за розміром пащі у самців цього виду очевидно дає змогу чинити опір у протистояннях з більшими за розмірами і сильнішими вужами та гадюками. Це формує у змій обережну реакцію до веретільниці. Самиці у порівнянні з самцями мають менші розміри голови і не здатні ефективно боронитись від змій. Але більш тонкі і довгі самиці веретільниці можуть проникати у вузенькі пустоти і нірки, куди не можуть протиснутись самці цього виду чи змії та В Поліссі веретільниця, як і інші види рептилій, недостатньо вивчені види. Ми ще дуже мало знаємо про біотопічне поширення цього виду, про його стосунки з іншими видами змій чи про загрози. Глобальне потепління у разі, коли у Поліссі не буде відбуватись зниження рівня ґрунтових вод та зникнення боліт очевидно не буде нести загроз цьому виду. Довготривалі спостереження у районі Поліського заповідника вказують навіть про зростання чисельності у деяких локалітетах та типах оселищ. Наукова література на українській чи російській мові у порівнянні з англомовною літературою містить обмежену інформацію по цьому виду.
Річний цикл життя
Шлюбний період у Поліссі проходить ймовірно у другій половині травня. Навесні самці часто б’ються за утримання індивідуальних територій чи для того, щоби відігнати суперників. Спаровування у цих спокійних і флегматичних ящірок може бути досить агресивним, коли самці міцно тримають самок, щільно їх затиснувши в щелепи. Турніри у цього виду можна побачити у місцях з порівняно високою чисельністю. У ставленні до людини веретільниці спокійні і майже не кусаються. Веретільниці довгожителі і доживають до 20 років і більше в дикій природі та понад 50 років - в неволі (Slow-worm…, 2006). Інтенсивність росту тіла найбільш висока у перший рік, коли ця тварина досягає довжини у 10 см. В наступні роки інтенсивність росту скорочується. Більшу частину року веретільниці проводять в укриттях. Часто дослідники для більш легкого пошуку цих тварин спеціально викладають металеві листи чи дошки, щоби під ними знаходити веретільниць. Вагітні самиці стають ще більш обережними. Народжує 7-14 (від 5 до 26) дитинчат. Період від запліднення до народження дитинчат складає біля 3 місяців. Довжина тіла новонароджених складає 5-6 см при вазі – 5,0-7,6 г. Линяє декілька разів на рік, залишаючи при цьому стару шкіру або виповзок (Пікулік, 1996).
Рис. 4. Шлюбний турнір між самцями. Один з суперників «помилково» кусає власне тіло (фото з Інтернету).

Статус у заповіднику
Нечисельний. Подекуди рідкісний. Обстежені площі пожеж 2021р. у ЧРЕБЗ не виявили фактів загибелі веретільниць або інших видів рептилій. Веретільниця веде прихований спосіб життя, складно виявляється у природі, особливо в умовах ЧРЕБЗ, тому вивчення поширення виду у різних типах оселищ потребує часу і встановлений статус цього виду є попереднім. В Білорусі чисельність веретільниць є нижчою чим в Українському Поліссі. Білоруські фахівці визнають факт низької чисельності цього виду та наводять такі показники чисельності: в цілому для лісових біогеоценозів вона становить 0,5 (від 0 до 50) особини на 1 га, в сосняках вона зустрічалася в 2 з 77 біотопів, в березняках - в 2 з 26, в вільшняках - в 3 з 52, а в ялинниках і дібровах взагалі вид не виявлений. При цьому щільність заселення склала в сосняку 0,02 особини на 1 га, у березниках - 0,4, вздовж узбіч доріг - 1,5, на заплавних луках -1,7 (Пикулик, 1988). Ці показники потрібно розглядати як умовні, бо щільності популяції веретільниці з точністю до десятих для всієї Білорусі наводити некоректно, бо на півдні і півночі цієї країни для одних і тих біотопів ці показники будуть різні. Окрім того, у межах одного й того ж лісового масиву щільність популяції веретільниці буде різною здавалось у одних і тих самих типах оселищ. Поширення рептилій у Поліссі є набагато мозаїчнішим і менш передбачуваним у порівнянні з дрібними мишоподібними чи птахами. Тому для певних типів оселищ доцільно наводити мінімальну, максимальну і середні щільності.
Література
Гащак С.П., Вишневський Д.О., Заліський О. О. Фауна хребетних тварин Чорнобильської зони відчуження (Україна). За заг. ред. С. П. Гащака. Славутич, 2006. 100 с.
Пікулік М. М. (ред.) . Земнаводныя. Паўзуны: Энцыклапедычны даведнік (Жывёльны свет Беларусі). Мiнск, 1996. 240с.
Пикулик М. М., Бахарев В. А., Косов С. В. "Пресмыкающиеся Белоруссии". Минск, 1988. 166с.
Федонюк О. В. Земноводні та плазуни заходу України. Львів: Сполом, 2006. 32 с.
Ananjeva, N.B., Orlov, N.L., Khalikov, R.G., Darevsky, I.S., Ryabov, S.A., Barabanov, A. (2006): The reptiles of northern Eurasia: taxonomic diversity, distribution, conservation status. Pentsoft Series Faunistica No: 47, Sofia-Moscow (Pensoft Publishers), 245 pp.
Badger, D., Netherton, J. (2003): Lizards: A natural history of some uncommon creatures, extraordinary chameleons, iguanas, geckos, and more. Voyageur Press, 160 pp.
Başoğlu, M., Baran, İ. (1977): Türkiye Sürüngenleri, kısım I, kaplumbağa ve kertenkeleler. 95 Ege Üniv., Fen Fakültesi, Kitaplar Serisi, No: 76,
Bornova, Izmir, Turkey. Carr, A.F. Jr. (1940): A contribution to the herpetology of Florida. University of Florida publication, Biological Science Series 3:1-118.
Claessen, D., de Roos, A.M., Persson, L. (2004): Population dynamic theory of size-dependent cannibalism. Proceedings the Royal Society Biological Science 271: 333-340.
Cooper, W.E. Jr. (1940): Prey odor discrimination by anguid lizards. Herpetologica 46(2): 183-190. Cooper, W.E. Jr., van Wyk, J.H. (1994): Absence of prey chemical discrimination by tongue-flicking in an ambush-foraging lizard having actively foraging ancestors. Ethology 97: 317-328. Cooper, W.F. Jr. (1997): Correlated evolution of prey chemical discrimination with foraging, lingual morphology and vomeronasal chemoreceptor abundance in lizards. Behavioral Ecology and Sociobiology 41: 257-265.
Davies, M. (1967): A case of Anguis fragilis devouring newly-born young. British Journal of Herpetology 4:20. Delitz, K. (1921): Kannibalismus bei einer blindschleiche. BL. Aquarienk. 32: 43-44.
Galán P., Ferreiro R. (2004). Reproductive ecology of the slow worm (Anguis fragilis) in the northwest Iberian Peninsula. In: Animal Biology. Online Publication Date: 01 Jan 2004. Volume 54: Issue 4. https://brill.com/view/journals/ab/54/4/article-p353_4.xml?language=en
Gvozˇdíka V., Jandzikc D., Lymberakisd P., Jablonskic D., Moravec J. (2010). Slow worm, Anguis fragilis (Reptilia: Anguidae) as a species complex: Geneticstructure reveals deep divergences. Molecular Phylogenetics and Evolution. 55 (2010). 460-472.
Fretey, J. (1987): Guide des Reptiles et Batraciens de France, Paris (Hatier). Ghira, I., Nemes, S., Rozsa, F. (1999): The ethnogram of Anguis fragilis: feeding behaviour. Nymphaea. Folia Naturae Bihariae XXVII: 153-159.
Hamilton, W.I.Jr., Pollack, J.A. (1961): The food of some lizards from Fort Benning, Georgia. Herpetologica 17: 99-106.
Luiselli L. (1992): The diet of the slow worm, Anguis fragilis f. Linnaeus, 1758, in the Tarvisio Forest (Carnic Alps, NE Italy). Herpetozoa 5: 91-94.
Luiselli, L., Capula, M., Anibaldi, C. (1994): Food habits of the slow worm, Anguis fragilis (L.), in two contrasting alpine environments. Bulletin de la Société Herpétologique de France 71/72: 45–48. Mitchell, J.C. (1986): Cannibalism in reptiles: a worldwide review. SSAR Herpetological Circulars 15: 1-37.
Mollov, I. (2010): A contribution to the knowledge of the trophic spectrum of the Slow Worm (Anguis fragilis L., 1758) (Reptilia: Anguidae) from Bulgaria. ZooNotes 9: 1-4.
Pedersen, I., Jensen, J., Toft, S. (2009): A method of obtaining dietary data for slow worms (Anguis fragilis) by means of non-harmful cooling and results from a Danish population. Journal of Natural History 43: 1011-1025.
Petzold H.G. (1971): Blindschleiche und Scheltopusik. Neue Brehm Bucherei, No. 448, Witenberg, Lutherstadi, GDR, 102 pp.
Polis, G.A. (1981): The evolution and dynamics of intraspecific predation. Annual Review of Ecology and Systematics 12: 225-251.
Polis, G.A., Myers, C.A. (1985): A survey of intraspecific predation among reptiles and amphibians. Journal of Herpetology 19: 99-107.
Tibor S., Gábor H. (2009). Sexual Size Dimorphism in Eastern Slow-Worm (Anguis fragilis colchica, Reptilia: Anguidae). Russian Qurnal Yerpetology, Vol 16, No 4 (2009).
The Herpetological Conservation Trust (The HCT) (2010): The Herpetological Conservation Trust slow worm fact sheet (October, 2010) http://www. herpconstrust.org.uk/animals/slow_worm.htm


 Чорнобильський радіаційно-екологічний біосферний заповідник
Tel: +38 (044) 275-01-88
Електронна пошта: info@zapovidnyk.org.ua
Чорнобильський радіаційно-екологічний біосферний заповідник
Tel: +38 (044) 275-01-88
Електронна пошта: info@zapovidnyk.org.ua







